The Diamondback Moth, Plutella xylostella (L.), is a notorious pest posing a substantial threat to global agriculture due to high resistance to conventional insecticides. Some P450 related enzymes are responsible to help the Diamondback Moth to metabolize and neutralize pesticides, contributing to the resistance problem. Our study aimed to understand the expression of the cytochrome P450 genes in response to Cypermethrin, Profenofos, and Acetamiprid. The P. xylostella larvae were collected from three different locations, representing varying resistance histories, and subjected them to controlled laboratory conditions for insecticide susceptibility and gene expression analysis. Expression levels of three genes –CYP321E1, CYP4M22, and CYP9G2- involved in the resistance in response to insecticide exposure were investigated. The analysis revealed significant variations in the gene expression patterns among the different populations and across the insecticides tested. The CYP4M22 gene displayed significant different expression patterns depending on the specific insecticide and population. The CYP321E1 and CYP9G2 genes also showed increased expression levels with prolonged exposure to the insecticides, indicating a potential adaptive mechanism for metabolizing pesticides. These findings emphasized the complex nature of insecticide resistance and suggested that gene expression patterns can differ significantly across populations, reflecting the unique evolutionary pressures in each environment.
| Published in | World Journal of Agricultural Science and Technology (Volume 2, Issue 4) |
| DOI | 10.11648/j.wjast.20240204.20 |
| Page(s) | 198-204 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Insecticide Resistance, Diamondback Moth, Pests, Cytochrome P450, Gene Expression
Gene | Sequence | m | Amplicon | Accession no. |
|---|---|---|---|---|
CYP321E1 | F: CTGGCTCACATTCTACGCAAC | 1 | 94 | KC626090 |
R: TGGCAGGTCTTATGATGAGGG | 1 | |||
CYP9G2 | F: TGGTGGGAAACTCTTTGACGA | 0 | 147 | AB096739 |
R: TGTTTTCGCGATGCTGCATAG | 0 | |||
CYP4M22 | F: CAGTGCAGGATAAAGTGGTGC | 0 | 111 | EU189050.1 |
R: CCTTGATACAGCACTCGAGGT | 0 | |||
Actin | F: CCGTGCCCATCTACGAAGGTTA | 128 | NM_001309126 | |
R: AGCGGTGGTGGTGAAGGAGTATC |
P450 | Cytochrome P450 |
CYP321E1 | Novel Cytochrome P450 Gene Number 321 |
CYP4M22 | Novel Cytochrome P450 Gene Number 4 |
CYP9G2 | Novel Cytochrome P450 Gene Number 9 |
LC50 | Lethal Concentration 50 |
EC | Emulsifiable Concentrate |
| [1] | Saran C, Genç HY, 2024. Genetic diversity of diamondback moth, Plutella xylostella L. (Lepidoptera: Plutellidae) populations in Türkiye. Molecular Biology Reports 51(1): 146. |
| [2] | Furlong MJ, Wright DJ, Dosdall LM, 2013. Diamondback moth ecology and management: problems, progress, and prospects. Annual Review of Entomology 58(1): 517-541. |
| [3] | Gul H, Gadratagi BG, Güncan A, et al., 2023. Fitness costs of resistance to insecticides in insects. Frontiers in Physiology 14: 1238111. |
| [4] | Santos VC, de Siqueira HA, da Silva JE, de Farias MJ, 2011. Insecticide resistance in populations of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), from the state of Pernambuco, Brazil. Neotropical Entomology 40(2): 264-70. |
| [5] | Wang J, Zheng X, Yuan J, et al., 2021. Insecticide Resistance Monitoring of the Diamondback Moth (Lepidoptera: Plutellidae) Populations in China. Journal of Economic Entomology 114(3): 1282-1290. |
| [6] | McGraw EA, O'neill SL, 2013. Beyond insecticides: new thinking on an ancient problem. Nature Reviews Microbiology 11(3): 181-193. |
| [7] | Zhang JJ, Yang H, 2021. Metabolism and detoxification of pesticides in plants. Science of the Total Environment 790: 148034. |
| [8] | Bahari A, Mehrzad J, Mahmoudi M, Bassami MR, Dehghani H, 2014. Cytochrome P450 isoforms are differently up-regulated in aflatoxin B(1)-exposed human lymphocytes and monocytes. Immunopharmacology & Immunotoxicology 36(1): 1-10. |
| [9] | Bahari A, Mehrzad J, Mahmoudi M, Bassami MR, Dehghani H, 2015. GST-M1 is transcribed moreso than AKR7A2 in AFB(1)-exposed human monocytes and lymphocytes. Journal of Immunotoxicology 12(2): 194-8. |
| [10] | Yu L, Tang W, He W, et al., 2015. Characterization and expression of the cytochrome P450 gene family in diamondback moth, Plutella xylostella (L.). Scientific Reports 5(1): 8952. |
| [11] | Baek JH, Clark JM, Lee SH, 2010. Cross-strain comparison of cypermethrin-induced cytochrome P450 transcription under different induction conditions in diamondback moth. Pesticide Biochemistry and Physiology 96(1): 43-50. |
| [12] | Li X, Li R, Zhu B, Gao X, Liang P, 2018. Overexpression of cytochrome P450 CYP6BG1 may contribute to chlorantraniliprole resistance in Plutella xylostella (L.). Pest Management Science 74(6): 1386-1393. |
| [13] | Nauen R, Zimmer CT, Andrews M, et al., 2012. Target-site resistance to pyrethroids in European populations of pollen beetle, Meligethes aeneus F. Pesticide Biochemistry and Physiology 103(3): 173-180. |
| [14] | Robertson JL, Preisler HK, 1992. Pesticide Bioassays W/ Arthropods. CRC-Press. |
| [15] | Schmittgen TD, Livak KJ, 2008. Analyzing real-time PCR data by the comparative C(T) method. Nature Protocols 3(6): 1101-1108. |
| [16] | Liu N, Li M, Gong Y, Liu F, Li T, 2015. Cytochrome P450s–Their expression, regulation, and role in insecticide resistance. Pesticide Biochemistry and Physiology 120: 77-81. |
| [17] | Finnigan JD, Young C, Cook DJ, Charnock SJ, Black GW, 2020. Cytochromes P450 (P450s): A review of the class system with a focus on prokaryotic P450s. Advances In Protein Chemistry and Structural Biology 122: 289-320. |
| [18] | Lin Q, Jin F, Hu Z, et al., 2013. Transcriptome analysis of chlorantraniliprole resistance development in the diamondback moth Plutella xylostella. PLoS One 8(8): e72314. |
| [19] | Villagra C, 2024. Role of epigenetic molecular mechanisms in insect evolutionary innovations. On Epigenetics and Evolution 1: 217-282. |
| [20] | Shen B, Zhao D, Qiao C, Lan W, 2004. Cloning of CYP9G2 from the diamondback moth, Plutella xylostella (Lepidoptera: Yponomeutidae). DNA Sequencing 15(3): 228-233. |
| [21] | Hu Z, Lin Q, Chen H, Li Z, Yin F, Feng X, 2014. Identification of a novel cytochrome P450 gene, CYP321E1 from the diamondback moth, Plutella xylostella (L.) and RNA interference to evaluate its role in chlorantraniliprole resistance. Bulletin of Entomological Research 104(6): 716-723. |
| [22] | Ninsin K, 2004. Selection for resistance to acetamiprid and various other insecticides in the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae). Journal of Applied Entomology 128(6): 445-451. |
| [23] | Khan S, Uddin MN, Rizwan M, et al., 2020. Mechanism of Insecticide Resistance in Insects/Pests. Polish Journal of Environmental Studies 29(3): 2023-2030. |
APA Style
Poodineh, M., Ravan, S., Sheikhigarjan, A., Sahebzade, N., Emamjome, A. (2024). Investigating Plutella xylostella Resistance to Insecticides: Sensitivity Shifts to Cypermethrin, Profenofos, and Acetamiprid. World Journal of Agricultural Science and Technology, 2(4), 198-204. https://doi.org/10.11648/j.wjast.20240204.20
ACS Style
Poodineh, M.; Ravan, S.; Sheikhigarjan, A.; Sahebzade, N.; Emamjome, A. Investigating Plutella xylostella Resistance to Insecticides: Sensitivity Shifts to Cypermethrin, Profenofos, and Acetamiprid. World J. Agric. Sci. Technol. 2024, 2(4), 198-204. doi: 10.11648/j.wjast.20240204.20
@article{10.11648/j.wjast.20240204.20,
author = {Mahdiyeh Poodineh and Sultan Ravan and Aziz Sheikhigarjan and Najme Sahebzade and Abbasali Emamjome},
title = {Investigating Plutella xylostella Resistance to Insecticides: Sensitivity Shifts to Cypermethrin, Profenofos, and Acetamiprid},
journal = {World Journal of Agricultural Science and Technology},
volume = {2},
number = {4},
pages = {198-204},
doi = {10.11648/j.wjast.20240204.20},
url = {https://doi.org/10.11648/j.wjast.20240204.20},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.wjast.20240204.20},
abstract = {The Diamondback Moth, Plutella xylostella (L.), is a notorious pest posing a substantial threat to global agriculture due to high resistance to conventional insecticides. Some P450 related enzymes are responsible to help the Diamondback Moth to metabolize and neutralize pesticides, contributing to the resistance problem. Our study aimed to understand the expression of the cytochrome P450 genes in response to Cypermethrin, Profenofos, and Acetamiprid. The P. xylostella larvae were collected from three different locations, representing varying resistance histories, and subjected them to controlled laboratory conditions for insecticide susceptibility and gene expression analysis. Expression levels of three genes –CYP321E1, CYP4M22, and CYP9G2- involved in the resistance in response to insecticide exposure were investigated. The analysis revealed significant variations in the gene expression patterns among the different populations and across the insecticides tested. The CYP4M22 gene displayed significant different expression patterns depending on the specific insecticide and population. The CYP321E1 and CYP9G2 genes also showed increased expression levels with prolonged exposure to the insecticides, indicating a potential adaptive mechanism for metabolizing pesticides. These findings emphasized the complex nature of insecticide resistance and suggested that gene expression patterns can differ significantly across populations, reflecting the unique evolutionary pressures in each environment.},
year = {2024}
}
TY - JOUR T1 - Investigating Plutella xylostella Resistance to Insecticides: Sensitivity Shifts to Cypermethrin, Profenofos, and Acetamiprid AU - Mahdiyeh Poodineh AU - Sultan Ravan AU - Aziz Sheikhigarjan AU - Najme Sahebzade AU - Abbasali Emamjome Y1 - 2024/12/30 PY - 2024 N1 - https://doi.org/10.11648/j.wjast.20240204.20 DO - 10.11648/j.wjast.20240204.20 T2 - World Journal of Agricultural Science and Technology JF - World Journal of Agricultural Science and Technology JO - World Journal of Agricultural Science and Technology SP - 198 EP - 204 PB - Science Publishing Group SN - 2994-7332 UR - https://doi.org/10.11648/j.wjast.20240204.20 AB - The Diamondback Moth, Plutella xylostella (L.), is a notorious pest posing a substantial threat to global agriculture due to high resistance to conventional insecticides. Some P450 related enzymes are responsible to help the Diamondback Moth to metabolize and neutralize pesticides, contributing to the resistance problem. Our study aimed to understand the expression of the cytochrome P450 genes in response to Cypermethrin, Profenofos, and Acetamiprid. The P. xylostella larvae were collected from three different locations, representing varying resistance histories, and subjected them to controlled laboratory conditions for insecticide susceptibility and gene expression analysis. Expression levels of three genes –CYP321E1, CYP4M22, and CYP9G2- involved in the resistance in response to insecticide exposure were investigated. The analysis revealed significant variations in the gene expression patterns among the different populations and across the insecticides tested. The CYP4M22 gene displayed significant different expression patterns depending on the specific insecticide and population. The CYP321E1 and CYP9G2 genes also showed increased expression levels with prolonged exposure to the insecticides, indicating a potential adaptive mechanism for metabolizing pesticides. These findings emphasized the complex nature of insecticide resistance and suggested that gene expression patterns can differ significantly across populations, reflecting the unique evolutionary pressures in each environment. VL - 2 IS - 4 ER -
Department of Plant Protection, Faculty of Agriculture, Zabol University, Zabol, Iran
Department of Plant Protection, Faculty of Agriculture, Zabol University, Zabol, Iran
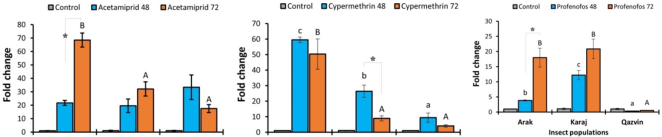
Figure 1. The expression levels (± SE) of the CYP321E1 gene in the three populations of Diamondback Moth in response to three insecticides at 48 h and 72 h challenge times. Control groups are included for each population to establish a baseline gene expression. Small and capital letters indicate significant differences at 48 h and 72 h sampling times between the populations, respectively.
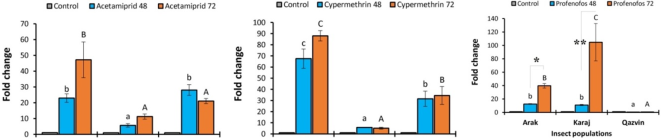
Figure 2. The expression levels (± SE) of the CYP4M22 gene in the three populations of Diamondback Moth in response to three insecticides at 48 h and 72 h challenge times. Control groups are included for each population to establish a baseline gene expression. Small and capital letters indicate significant differences at 48 h and 72 h sampling times between the populations, respectively.
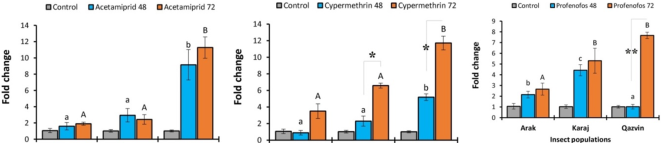
Figure 3. The expression levels (± SE) of the CYP4M22 gene in the three populations of Diamondback Moth in response to three insecticides at 48 h and 72 h challenge times. Control groups are included for each population to establish a baseline gene expression. Small and capital letters indicate significant differences at 48 h and 72 h sampling times between the populations, respectively.
Information

